Cells are divided into several compartments, each with a characteristic structure,biochemical composition, and function (see illustration). These compartments arecalled organelles. They are delimited by membranes composed of phospholipidbilayers and a number of proteins specialized for each type of organelle. Alleukaryotic cells have a nucleus surrounded by a nuclear envelope, and a plasmamembrane that borders the whole cell. Most eukaryotic cells also haveendoplasmic reticulum, a Golgi apparatus, lysosomes, mitochondria, andperoxisomes. Plant cells have chloroplasts for photosynthesis in addition to theorganelles that both they and animal cells possess. These organelles aresuspended in a gellike cytoplasmic matrix composed of three types of proteinpolymers called actin filaments, microtubules, and intermediate filaments. Inaddition to holding the cell together, the actin filaments and microtubules act astracks for several different types of motor proteins that are responsible for cellmotility and organelle movements within the cytoplasm.
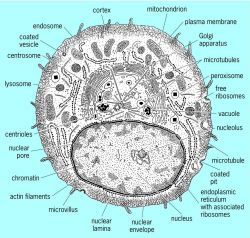
Section through an animal cell showing the majorcomponents visible by electron microscopy
A major challenge in the field of cell biology is to learn how each organelle andthe cytoplasmic matrix are assembled and distributed in the cytoplasm. This is avery complex process since cells consist of more than 2000 different proteinmolecules together with a large number of lipids, polysaccharides, and nucleicacids, including both deoxyribonucleic acid (DNA) and many different types ofribonucleic acid (RNA). See Nucleic acid
The cell must possess enough information to specify which molecules are to beassociated in a specific compartment, to route the appropriate groups ofmolecules to their compartments, and then to position each type of componentappropriately in the cell. As a result of intense research on each of these topics, anumber of specific chemical reactions that contribute to organizing cells are nowrecognized, but even more important, a small number of general principles thatexplain these complex processes of life can be appreciated.
Normal cells regulate the production and degradation of all of their constituentmolecules so that the right balance of molecules is present at any given time. Thegenes stored in nuclear DNA are duplicated precisely once per cell cycle. Thesupply of each of thousands of proteins is usually regulated at the level of thegenes at the time of biosynthesis and by the rate of degradation. These proteinsserve as enzymes that determine the rate of synthesis of themselves as well as ofother cellular components such as nucleic acids, carbohydrates, and lipids. Eachof these processes is regulated by molecular feedback loops to assure the properlevels of each cellular constituent. See Cell cycle, Cell metabolism
A large majority of cellular components are generated by the self-assembly oftheir constituent molecules. Self-assembly means that the information required formolecules to bind together in the proper orientation is contained in the moleculesthemselves. Some examples include the binding of histones to DNA, theformation of bilayers from phospholipids, and the polymerization of actinmolecules into filaments. The molecules are usually brought together by diffusion.The energy required to hold them together derives from the exclusion of waterfrom their complementary surfaces as well as from the formation of ionic bondsand hydrogen bonds. The variety of molecular structures found in proteins allowseach type to self-assemble specifically, only with their correct partner molecules.
After biosynthesis, proteins and nucleic acids are routed to their proper cellularcompartment by specific recognition signals consisting of parts of the molecule or,in the case of some proteins, by sugar side chains. These signals are recognizedby compartment-specific receptors that guide the molecules to the correctcompartment. For example, proteins destined for lysosomes have a specificsugar side chain added in the Golgi apparatus that guides them to lysosomes.Similarly, proteins destined for the nucleus all contain short sequences of aminoacids that target the proteins for uptake by the nucleus. These so-called nuclearrecognition sequences most likely bind to specific receptors associated with thenuclear pores, the channels through the nuclear envelope that connect thenucleus with the cytoplasm.
Most molecules move to their correct compartment by the process of diffusiondown concentration gradients, but organelles generally require transport systemscomposed of microtubules or actin filaments together with specific motor proteinsto position them correctly in the cytoplasm. For example, a protein moleculedestined to be part of a mitochondrion will diffuse from the site of biosynthesisthrough the cytoplasm to a mitochondrion, where it will bind to a receptor thatguides its incorporation into the mitochondrion. On the other hand, themitochondrion itself is too large to diffuse through the network of protein fibers inthe cytoplasmic matrix, and so it must be pulled through the matrix by a motorprotein that moves along microtubules to the correct place in the cell.